400-998-5282
专注多肽 服务科研
400-998-5282
专注多肽 服务科研
Preptin is a 34-amino acid peptide hormone, corresponding to Asp⁶⁹-Leu¹⁰² of the proinsulin-like growth factor II E-peptide. It is co-secreted with insulin and amylin from the pancreatic β-cells and is a physiological amplifier of glucose-m
编号:198851
CAS号:315197-69-4
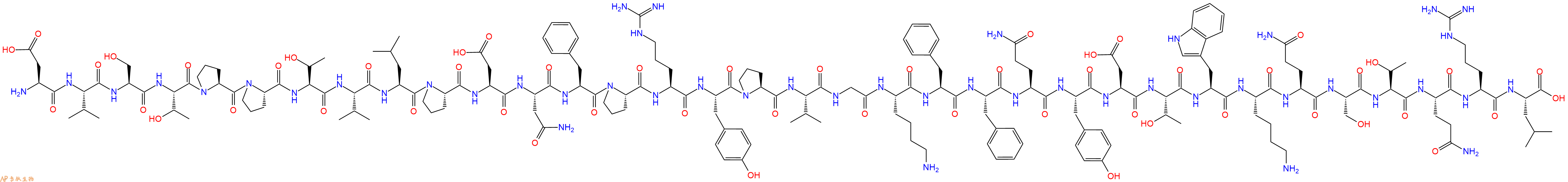
单字母:H2N-DVSTPPTVLPDNFPRYPVGKFFQYDTWKQSTQRL-OH
Preptin is a 34-amino acid peptide hormone, corresponding to Asp⁶⁹-Leu¹⁰² of the proinsulin-like growth factor II E-peptide. It is co-secreted with insulin and amylin from the pancreatic β-cells and is a physiological amplifier of glucose-mediated insulin secretion. It is also anabolic to bone tissue and may contribute to the preservation of bone mass observed in hyperinsulinemic states such as obesity.
重组血小板衍生生长因子 AB(异源二聚体)(人)形成混合链二聚体,由胱氨酸结相关稳定化作用。序列排列支持受体结合不对称性。折叠动力学影响配体取向与信号启动。应用包括生长因子受体建模与异源二聚体结构研究。
rec PDGF AB (heterodimer) (human) forms a mixed-chain dimer with cysteine knot–related stabilization. Sequence arrangement supports receptor-binding asymmetry. Folding dynamics influence ligand orientation and signaling initiation. Applications include growth-factor receptor modeling and heterodimer structural studies.
Preptin (human)是一个对应的是胰岛素样生长因子II E肽(proinsulin-like growth factor II E-peptide) Asp69-Leu102残基序列的34个氨基酸的多肽类激素。它在胰岛β细胞中与胰岛素和amylin一起被共分泌出来,是葡萄糖调节胰岛素分泌的生理放大器。它同时也参与骨组织的新陈代谢,在高胰岛素状态比如肥胖中观察到Preptin (human)有可能对保护骨量有帮助。
胰岛素样生长因子(Insulin-Like Growth Factors, IGF)的定义
胰岛素样生长因子(IGF)-1和IGF-2是普遍表达的肽类,其序列与胰岛素具有同源性。
Insulin-like growth factors (IGF)-1 and IGF-2 are ubiquitously expressed peptides with sequence homology to insulin.
胰岛素样生长因子(Insulin-Like Growth Factors, IGF)的相关多肽
胰岛素样生长因子(IGFs)与细胞膜上的特定受体,即胰岛素样生长因子-I受体(IGF-IR)相互作用,且这种相互作用受一组特异性结合蛋白的调节。所有这些分子均被视为胰岛素样生长因子(IGF)家族的成员,该家族包括多肽配体胰岛素样生长因子-I(IGF-I)和胰岛素样生长因子-II(IGF-II)、两种细胞膜受体(即IGF-IR和IGF-IIR)以及六种胰岛素样生长因子结合蛋白(即IGFBP-1至IGFBP-6)。
IGFs interacts with a specific receptor on the cell membrane, namely, the IGF-I receptor (IGF-IR), and the interaction is regulated by a group of specific binding proteins. All of these molecules are considered to be members of the IGF family, which includes the polypeptide ligands IGF-I and IGF-II, two types of cell membrane receptors (i.e., IGF-IR and IGF-IIR), and six IGF-binding proteins (i.e., IGFBP-1 through IGFBP-6).
胰岛素样生长因子(Insulin-Like Growth Factors, IGF)的结构特征
IGFs
胰岛素样生长因子-I(IGF-I)和胰岛素样生长因子-II(IGF-II)是单链多肽。这两种分子的氨基酸序列有62%的同源性。它们在结构上还有其他相似之处,且结构类似于胰岛素原【1】。IGF-I由70个氨基酸残基组成,IGF-II由67个氨基酸残基组成,分为A和B域(类似于胰岛素)、C域(类似于胰岛素原的连接肽)和D域(胰岛素中不存在)。IGF-I和IGF-II中的三个链内二硫键已被证明位于与(胰岛素)原胰岛素中类似的位置【1】。
IGF-I and IGF-II are single-chain polypeptides. The two molecules have 62% homology in their amino acid sequences. The molecules share additional structural similarities, and their structures resemble the structure of proinsulin【1】. IGF I consists of 70 amino acid residues, IGF I1 of 67, grouped into domains A and B (similar to insulin), C (analogous to the connecting peptide of proinsulin) and D (not present in insulins). The three intrachain disulfide bridges in IGF 1 and I1 have shown to be located in analogous positions to those in (pro) insulin【1】.
IGFBPs
哺乳动物胰岛素样生长因子结合蛋白(IGFBPs)的一级结构似乎包含三个大小大致相当的不同结构域:保守的N端结构域、高度可变的中区结构域和保守的C端结构域。N端结构域在信号肽后包含80-93个氨基酸残基。前体肽中的16-20个半胱氨酸中有10-12个位于该结构域内。在IGFBP-1至-5中,这12个半胱氨酸完全保守,而在IGFBP-6中,12个半胱氨酸中有10个是不变的【2】。中区结构域的大小从55个氨基酸残基到95个氨基酸不等,将N端结构域与C端结构域分隔开。每个中区片段的氨基酸序列似乎对该蛋白质是独特的。C端结构域高度保守,在总共16-20个半胱氨酸中,有6个位于C端,并且是严格保守的【2】。
The primary structures of mammalian IGFBPs appear to contain three distinct domains of roughly equivalent sizes: the conserved N-terminal domain, the highly variable midregion, and the conserved C-terminal domain.N-terminal domain contains 80–93 amino acid residues after the signal. Ten to 12 of the 16–20 cysteines found in the prepeptides are located within this domain. In IGFBP-1 to -5, these 12 cysteines are fully conserved, whereas in IGFBP-6, 10 of the 12 cysteines are invariant【2】.Midregion ranging in size from 55 amino acid residues to 95 amino acids separates the N-terminal domain from the C-terminal domain. The amino acid sequence for each midsegment appears to be unique to the protein. C-terminal region are highly conserved and, 6 cysteines of the total 16–20 cysteines are found in the C terminus and are strictly conserved【2】.
IGF Receptors
胰岛素样生长因子受体I(IGF-IR)和胰岛素样生长因子受体II(IGF-IIR)都是糖蛋白,位于细胞膜上。IGF-IR是由两个相同的α亚基和两个相同的β亚基组成的四聚体。从结构上看,IGF-IR与胰岛素受体相似,两者之间有60%的同源性。IGF-IIR是单聚体。受体的细胞外结构域中有三个配体结合区,一个用于结合IGF-II,两个用于结合含有甘露糖-6-磷酸(M6P)的蛋白质,包括肾素、增殖素、甲状腺球蛋白和(转化生长因子β(TGF-β)的潜在形式【2】。
Both IGF-IR and IGF-IIR are glycoproteins and are located on the cell membrane. IGF-IR is a tetramer of two identical a-subunits and two identical ß-subunits. Structurally, IGF-IR resembles the insulin receptor, and there is 60% homology between them. IGF-IIR is monomeric. Three ligand-binding regions are found in the extracellular domain of the receptor, one for IGF-II binding and two for proteins containing mannose-6-phosphate (M6P), including renin, proliferin, thyroglobulin, and the latent form of (TGF)-ß transforming growth factor【2】.
胰岛素样生长因子(Insulin-Like Growth Factors, IGF)的作用机制
胰岛素样生长因子(IGFs)与胰岛素样生长因子受体(IGF-IR)结合会激活受体的酪氨酸激酶活性,从而引发一系列级联反应。目前已经确定了IGF-IR的两条不同信号转导通路。一条通路激活Ras蛋白、Raf蛋白和丝裂原活化蛋白激酶,另一条通路则涉及磷酸肌醇-3-激酶。IGF-IR参与细胞转化。体外实验表明,通过消除IGF-IR基因、抑制其表达或抑制其功能,从而从细胞膜上移除IGF-IR,可以消除细胞转化【3】。胰岛素样生长因子结合蛋白(IGFBPs)具有多种复杂功能。IGFBPs能够抑制或增强IGFs的作用,从而抑制或刺激细胞增殖。IGFBPs对IGFs的这些相反作用由结合蛋白的分子结构决定。当与IGFs结合时,IGFBPs发挥三个主要作用:1)运输IGFs,2)保护IGFs免受降解,3)调节IGFs与IGF-IR之间的相互作用。通常,IGFBPs对IGFs的结合亲和力高于IGF-IR;因此,IGFBPs与IGFs的结合会阻断IGFs与IGF-IR之间的相互作用,从而抑制IGF的作用。然而,IGFBPs与IGFs的结合也能保护IGFs免受蛋白酶降解,这种保护作用可通过增加IGFs在局部组织中的生物利用度来增强其作用【3】。
Binding of IGFs to IGF-IR activates the receptor's tyrosine kinase activity, which triggers a cascade of reactions. Two distinct signal transduction pathways have been identified for IGF-IR. One pathway activates Ras protein, Raf protein, and mitogen-activated protein kinase, and the other pathway involves phosphoinositol-3-kinase. IGF-IR is involved in cell transformation. In vitro experiments have shown that removal of IGF-IR from the cell membrane by eliminating the IGF-IR gene, by suppressing its expression, or by inhibiting its function can abolish cell transformation【3】. IGFBPs have multiple and complex functions. IGFBPs are able to inhibit or to enhance the action of IGFs, resulting in either suppression or stimulation of cell proliferation. These opposing effects of IGFBPs on IGFs are determined by the molecular structures of the binding proteins. When binding to IGFs, IGFBPs play three major roles: 1) transporting IGFs, 2) protecting IGFs from degradation, and 3) regulating the interaction between IGFs and IGF-IR. Normally, IGFBPs have higher binding affinity to IGFs than does IGF-IR; therefore, binding of IGFBPs to IGFs blocks the interaction between IGFs and IGF-IR and suppresses IGF action. However, binding of IGFBPs to IGFs also protects IGFs from proteolytic degradation, and that protection can enhance the action of IGFs by increasing their bioavailability in local tissue【3】.
胰岛素样生长因子(Insulin-Like Growth Factors, IGF)的功能
直接参与癌症——胰岛素样生长因子-I(IGF-I)和胰岛素样生长因子-II(IGF-II)是多种癌细胞系的强促有丝分裂剂。动物实验表明,IGF-I的过表达会增加某些组织中肿瘤发展的可能性。胰岛素样生长因子(IGFs)对癌细胞的作用是通过胰岛素样生长因子受体(IGF-IR)介导的。从细胞膜中消除IGF-IR、阻断IGFs与IGF-IR的相互作用或中断IGF-IR的信号转导通路,均可消除IGFs对癌细胞的促有丝分裂作用。IGF-IR在某些癌症中过表达,且其过表达与侵袭性肿瘤相关。最近的一项研究表明,胰岛素受体参与介导IGF-II对乳腺癌的作用。具有强转移倾向的癌细胞比转移能力低的癌细胞具有更高的IGF-II和IGF-IR表达。在癌症中,胰岛素样生长因子结合蛋白(IGFBPs)调节IGFs的作用。在大多数情况下,结合蛋白会抑制IGFs的促有丝分裂作用并促进细胞凋亡。研究表明,IGFBP-3在不与IGFs相互作用的情况下抑制了乳腺癌细胞的生长【4】。
Direct Involvement in Cancer - IGF-I and IGF-II are strong mitogens for a wide variety of cancer cell lines. Animal experiments indicate that overexpression of IGF-I increase the likelihood of tumor development in certain tissues. The effects of IGFs on cancer cells are mediated through IGF-IR. Eliminating IGF-IR from the cell membrane, blocking the interaction of IGFs with IGF-IR, or interrupting the signal transduction pathway of IGF-IR can abolish the mitogenic action of IGFs on cancer cells. IGF-IR is overexpressed in certain cancers, and its overexpression is associated with aggressive tumors. A recent study indicates that the insulin receptor is involved in mediating the actions of IGF-II on breast cancer. Cancer cells with a strong tendency to metastasize have higher expression of IGF-II and IGF-IR than those with a low ability to do so.In cancer, IGFBPs regulate the action of IGFs. In most situations, the binding proteins suppress the mitogenic action of IGFs and promote apoptosis. It has been shown that IGFBP-3 inhibited breast cancer cell growth without interacting with IGFs【4】.
胰岛素样生长因子I(IGF-I)保护并拯救海马神经元免受β-淀粉样蛋白和人类胰淀素诱导的毒性影响——胰岛素样生长因子(IGF-I和IGF-II)是众所周知的营养因子,其特异性受体在大脑中分布独特,尤其集中在海马结构中。IGF在海马区具有神经营养活性,而海马区是阿尔茨海默病中受严重影响的一个区域。有证据表明,β-淀粉样蛋白(aβ)衍生的肽可能在阿尔茨海默病中观察到的神经退行性过程中发挥重要作用,已有研究表明,IGF可对海马神经元提供神经保护,使其免受淀粉样蛋白衍生物的毒性影响【5】。
IGF I protects and rescues hippocampal neurons against ß-amyloid- and human amylin-induced toxicity - Insulin-like growth factors (IGF-I and IGF-II) are well known trophic factors and their specific receptors are uniquely distributed throughout the brain, being especially concentrated in the hippocampal formation. IGFs possess neurotrophic activities in the hippocampus, an area severely affected in Alzheimer disease. There is evidence that ß-amyloid (aß)-derived peptides likely play an important role in the neurodegenerative process observed in Alzheimer disease, it has been shown that IGFs can be neuroprotective to hippocampal neurons against toxicity induced by amyloidogenic derivatives【5】.
胰岛素样生长因子(Insulin-Like Growth Factors, IGF)的相关文献
1. Daughaday WH, Rotwein P (1989). Insulin-like growth factors I and II - Peptide, messenger ribonucleic acid and gene structures, serum, and tissue concentrations. Endocr. Rev, 10:68–91.
2. Jones JI, Clemmons DR (1995). Insulin-like growth factors and their binding proteins: biological actions. Endocr. Rev., 16:3–34.
3. Clemmons DR (1997). Insulin-like growth factor binding proteins and their role in controlling IGF actions. Cytokine Growth Factor Rev, 8:45–62.
4. Yu H, Rohan T (2000). Role of the Insulin-Like Growth Factor Family in Cancer Development and Progression. Journal of the National Cancer Institute., 92 (18):1472-1489.
5. Doré S, Kar S, Quirion R (1997). Insulin-like growth factor I protects and rescues hippocampal neurons against ß-amyloid- and human amylin-induced toxicity. Proc. Natl. Acad. Sci, 94:4772–4777.
| DOI | 名称 | |
|---|---|---|
| 10.1042/0264-6021:3600431 | Preptin derived from proinsulin-like growth factor II (proIGF-II) is secreted from pancreatic islet beta-cells and enhances insulin secretion | 下载 |
| 10.1152/ajpendo.00642.2005 | Preptin, another peptide product of the pancreatic beta-cell, is osteogenic in vitro and in vivo | 下载 |
| 10.1080/07853890802244142 | Circulating preptin levels in normal, impaired glucose tolerance, and type 2 diabetic subjects | 下载 |
多肽H2N-Asp-Val-Ser-Thr-Pro-Pro-Thr-Val-Leu-Pro-Asp-Asn-Phe-Pro-Arg-Tyr-Pro-Val-Gly-Lys-Phe-Phe-Gln-Tyr-Asp-Thr-Trp-Lys-Gln-Ser-Thr-Gln-Arg-Leu-COOH的合成步骤:
1、合成CTC树脂:称取2.43g CTC Resin(如初始取代度约为0.73mmol/g)和2.13mmol Fmoc-Leu-OH于反应器中,加入适量DCM溶解氨基酸(需要注意,此时CTC树脂体积会增大好几倍,避免DCM溶液过少),再加入5.32mmol DIPEA(Mw:129.1,d:0.740g/ml),反应2-3小时后,可不抽滤溶液,直接加入1ml的HPLC级甲醇,封端半小时。依次用DMF洗涤2次,甲醇洗涤1次,DCM洗涤一次,甲醇洗涤一次,DCM洗涤一次,DMF洗涤2次(这里使用甲醇和DCM交替洗涤,是为了更好地去除其他溶质,有利于后续反应)。得到 Fmoc-Leu-CTC Resin。结构图如下:

2、脱Fmoc:加3倍树脂体积的20%Pip/DMF溶液,鼓氮气30分钟,然后2倍树脂体积的DMF 洗涤5次。得到 H2N-Leu-CTC Resin 。(此步骤脱除Fmoc基团,茚三酮检测为蓝色,Pip为哌啶)。结构图如下:

3、缩合:取5.32mmol Fmoc-Arg(Pbf)-OH 氨基酸,加入到上述树脂里,加适当DMF溶解氨基酸,再依次加入10.64mmol DIPEA,5.06mmol HBTU。反应30分钟后,取小样洗涤,茚三酮检测为无色。用2倍树脂体积的DMF 洗涤3次树脂。(洗涤树脂,去掉残留溶剂,为下一步反应做准备)。得到Fmoc-Arg(Pbf)-Leu-CTC Resin。氨基酸:DIPEA:HBTU:树脂=3:6:2.85:1(摩尔比)。结构图如下:

4、依次循环步骤二、步骤三,依次得到
H2N-Arg(Pbf)-Leu-CTC Resin
Fmoc-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Pro-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Pro-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Pro-Pro-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Pro-Pro-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Thr(tBu)-Pro-Pro-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Thr(tBu)-Pro-Pro-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Ser(tBu)-Thr(tBu)-Pro-Pro-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Ser(tBu)-Thr(tBu)-Pro-Pro-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Val-Ser(tBu)-Thr(tBu)-Pro-Pro-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
H2N-Val-Ser(tBu)-Thr(tBu)-Pro-Pro-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
Fmoc-Asp(OtBu)-Val-Ser(tBu)-Thr(tBu)-Pro-Pro-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin
以上中间结构,均可在专肽生物多肽计算器-多肽结构计算器中,一键画出。
最后再经过步骤二得到 H2N-Asp(OtBu)-Val-Ser(tBu)-Thr(tBu)-Pro-Pro-Thr(tBu)-Val-Leu-Pro-Asp(OtBu)-Asn(Trt)-Phe-Pro-Arg(Pbf)-Tyr(tBu)-Pro-Val-Gly-Lys(Boc)-Phe-Phe-Gln(Trt)-Tyr(tBu)-Asp(OtBu)-Thr(tBu)-Trp(Boc)-Lys(Boc)-Gln(Trt)-Ser(tBu)-Thr(tBu)-Gln(Trt)-Arg(Pbf)-Leu-CTC Resin,结构如下:
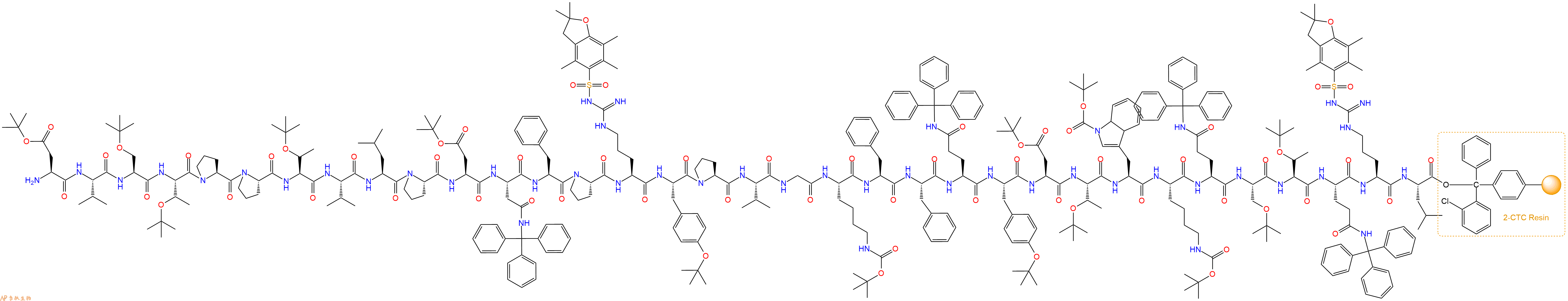
5、切割:6倍树脂体积的切割液(或每1g树脂加8ml左右的切割液),摇床摇晃 2小时,过滤掉树脂,用冰无水乙醚沉淀滤液,并用冰无水乙醚洗涤沉淀物3次,最后将沉淀物放真空干燥釜中,常温干燥24小试,得到粗品H2N-Asp-Val-Ser-Thr-Pro-Pro-Thr-Val-Leu-Pro-Asp-Asn-Phe-Pro-Arg-Tyr-Pro-Val-Gly-Lys-Phe-Phe-Gln-Tyr-Asp-Thr-Trp-Lys-Gln-Ser-Thr-Gln-Arg-Leu-COOH。结构图见产品结构图。
切割液选择:1)TFA:H2O=95%:5%
2)TFA:H2O:TIS=95%:2.5%:2.5%
3)三氟乙酸:茴香硫醚:1,2-乙二硫醇:苯酚:水=87.5%:5%:2.5%:2.5%:2.5%
(前两种适合没有容易氧化的氨基酸,例如Trp、Cys、Met。第三种适合几乎所有的序列。)
6、纯化冻干:使用液相色谱纯化,收集目标峰液体,进行冻干,获得蓬松的粉末状固体多肽。不过这时要取小样复测下纯度 是否目标纯度。
7、最后总结:
杭州专肽生物技术有限公司(ALLPEPTIDE https://www.allpeptide.com)主营定制多肽合成业务,提供各类长肽,短肽,环肽,提供各类修饰肽,如:荧光标记修饰(CY3、CY5、CY5.5、CY7、FAM、FITC、Rhodamine B、TAMRA等),功能基团修饰肽(叠氮、炔基、DBCO、DOTA、NOTA等),同位素标记肽(N15、C13),订书肽(Stapled Peptide),脂肪酸修饰肽(Pal、Myr、Ste),磷酸化修饰肽(P-Ser、P-Thr、P-Tyr),环肽(酰胺键环肽、一对或者多对二硫键环),生物素标记肽,PEG修饰肽,甲基化修饰肽等。
以上所有内容,为专肽生物原创内容,请勿发布到其他网站上。