400-998-5282
专注多肽 服务科研
400-998-5282
专注多肽 服务科研
核糖体合成的阳离子抗菌肽或宿主防御肽通过与细菌表面带负电荷脂质的静电相互作用显示出了广谱抗菌活性。与正常细胞相比,由于癌细胞表面的磷脂酰丝氨酸(带负电荷)比例增加,因此阳离子双亲肽可能是抗肿瘤药物的一个有效治疗方法,并且具有高选择性。抗菌肽的抗肿瘤机制从作用模式上可以分为2 类:选择性膜破坏和非膜溶解作用机制,其代表性的抗肿瘤肽包括α防御素1、乳铁蛋白 B 等。抗菌肽抗肿瘤的临床应用主要制约在其稳定性及给药途径,通过对其进行结构优化,开发创新配方和药物输送系统,使这些抗菌肽在未来肿瘤治疗过程中发挥更大的作用。
研究发现新的天然肿瘤杀伤分子促使抗肿瘤药物的更新不断发展,这其中包括抗菌肽这类具有多种性能 生物活性的分子11-21。目前的抗肿瘤治疗以手术治疗、化学治疗及放射治疗为主,这些治疗方法在延长肿瘤患者 生存时间方面仍起着至关重要的作用3-4]。肿瘤细胞是一 类快速分裂增殖的细胞,其生长过程并不受正常生长过程调控;化学治疗的主要目的是干扰这类非正常细胞的 分裂过程同。然而,很多抗肿瘤药物缺乏特异性,在其 抗肿瘤过程中对快速增殖的正常细胞亦有杀伤作用,同时容易使肿瘤细胞产生抗药性同。因此,不断开发新型 抗肿瘤药物来控制这些非正常细胞分裂是很有必要的。 其中,抗菌肽作为一种可从自然中提取并用于对抗肿瘤 细胞的潜在候选药物,引起了科学家们广泛关注。抗菌 肽是 一 类产生低抗药性并对快速增殖细胞产生毒性的宿主免疫基本成分,其主要作用基团是带正电的氨基酸 片段16-7。文章综述人类细胞源性抗菌肽的抗肿瘤相关作 用机制,阐述抗菌肽在肿瘤治疗中的应用现状。
2 结果 Results
2.1 抗菌肽的概述 抗菌肽存在于所有形式的生命体 中,包括原核生物、真菌、植物和人类图。尽管相同的 多肽序列很少出现在不同的物种中,但抗菌肽仍具有一些共同的结构特征。抗菌肽具有阳离子和两亲性结构, 可以通过静电和疏水作用选择性地与带负电荷的微生 物膜结合,从而导致膜破裂或移位。因此,微生物膜通 常被认为是抗菌肽的主要作用靶点,并根据这种认知提 出了几种抗菌活性机制9。抗菌肽的抗菌机制从作用模 式上可以分为2类主要的模式:直接致死模式和免疫调 节模式,直接致死作用机制可进一步分为膜靶向和非膜 靶向机制[10]。而发现抗菌肽抗菌活性的同时,其抗肿瘤 机制也被逐渐阐明。
2.2 抗菌肽的性质
2.2.1 抗菌肽的生物转化及理化特性 抗菌肽是 一 类 特殊的分子,通常具有5-40位氨基酸残基,带有正电荷 且具有两亲结构[1];抗菌肽通过静电相互作用与带有负 电的细胞膜脂质相互识别,进而作用于靶点之上[12-13]。 在人体中抗菌肽可以在很多组织中表达,包括皮肤上皮 细胞、呼吸系统、消化系统甚至是免疫系统中,见图2。根据合成位点的不同,抗菌肽又可分为非核糖体肽和核 糖体肽,前者一般在细菌和真菌的胞质中合成,后者广 泛合成于原核和真核细胞的核糖体中[15。抗菌肽在结构 上是不均匀的,可能呈线性(具有两亲性和疏水性的α - 螺旋残基)、β-折叠、环状、脂质、大环或α-螺旋杆构 象。值得注意的是这种多样性结构来自翻译后修饰, 包括糖基化、磷酸化和酰胺化等16。抗菌肽对于很多微生物均有效果,其中包括细菌、 真菌及病毒。值得注意的是,内源性抗菌肽被认为对病 原体具有高度的选择性。介导其抗菌作用的经典机制包 括以下几方面:穿透质膜或细胞壁,从而导致离子梯度的溶解或破坏;与核酸连接并破坏其功能;破坏保持微 生物细胞壁完整性的酶的功能[17。

2.2.2 抗菌肽的免疫调节作用 抗菌肽除了在执行杀 菌作用方面的典型功能外,亦可与宿主细胞相互作用。例如β-防御素, 一种对革兰阳性菌、革兰阴性菌及病毒 都有作用的抗菌肽,同样也是CCR6 趋化因子受体的配 体,可在T淋巴细胞和树突状细胞上表达;因此它能够 作为宿主细胞主动与被动免疫的桥梁。此外,抗菌肽还 对其他宿主免疫细胞产生影响:包括改变宿主基因表 达,诱导趋化因子分泌,调节中性粒细胞, T 淋巴细胞 和树突状细胞的激活或死亡,以及调节细胞分化途径, 促进免疫介导的伤口愈合等作用[18-19。但上述的抗菌肽 与宿主细胞之间的相互作用的具体机制迄今为止尚未 阐明。
2.3 抗菌肽对抗肿瘤细胞的特性
2.3.1 抗菌肽的基本抗肿瘤特性 众所周知,革兰阴性 细菌外膜上的脂多糖和革兰阳性细菌膜上的脂磷烯酸 导致细菌表面上具有高密度的负电荷l²0-211。如前所述, 抗菌肽表面带有正电荷,因此抗菌肽与细菌细胞膜具有 很强的相互作用。值得注意的是,由于具有各种类型二 级结构的阳离子抗菌肽(典型的携带电荷为+2或更多)和 具有不同正电荷数的序列,均能够识别其微生物靶点, 因此并没有一致的活性结合序列[22]。因此,阳离子两性 基团是抗菌肽具有活性的主要特征,而氨基酸序列决定 其活性特异性。虽然抗菌肽可能对哺乳动物细胞存在毒 性,但毒性浓度通常比对细菌的最低抑菌浓度高一个对 数数量级l23。因此,合理地预测抗菌肽的最小治疗指数 (最大耐受剂量/最小治疗剂量)是其抗肿瘤治疗的前提。
磷脂酰丝氨酸(占磷脂总量的3%-9%)通常存在于 正常哺乳动物细胞胞质膜的内叶上。肿瘤细胞中磷脂 酰丝氨酸表达转移至质膜的外叶中,这使得一些特殊 的抗菌肽能够与之特异性靶定,见图 3 。当细胞发生凋 亡时,磷脂酰丝氨酸可以被转移到质膜的外叶,从而 破坏哺乳动物正常细胞膜的不对称性125。这种不对称 性的改变通常存在于凋亡细胞和一些肿瘤细胞中,进 而有助于其被单核细胞识别而被其清除。其他可能导致肿瘤细胞负电荷升高的因素包括硫酸肝素和肿瘤细 胞表面存在O- 糖基化黏蛋白[26]。然而,与细菌细胞膜 上的负电荷相比,癌细胞上的负电荷密度相对较低, 因此,抗菌肽对于癌细胞的亲和力弱于细菌。与正常 细胞膜相比,肿瘤细胞膜存在抗菌肽的潜在靶点。一 些肿瘤细胞因细胞膜中较低水平的胆固醇而导致细胞 膜流动性增加。例如与正常白细胞和肺细胞相比,在 白血病和肺癌患者中,其肿瘤细胞的细胞膜由于其膜 中胆固醇水平较低而显示出其流动性增加127],从而最 终增强了抗菌肽的溶解作用[28]。肿瘤细胞另一个特性 是其微绒毛数量增加,进而使细胞膜表面积随之增大, 最终增强抗菌肽与肿瘤细胞的结合能力l²4。一 旦与癌 细胞结合,抗菌肽可能破坏细胞膜或穿透细胞,攻击 线粒体,导致细胞凋亡。防御素(长29-45个氨基酸)是 一类重要的富含半胱氨酸的抗菌肽,防御素a 的抗肿瘤 活性,特别是人类中性粒细胞肽1-3,已被证实可以通 过膜溶解和促进凋亡及抑制血管生成等作用而发挥抗 肿瘤活性l2]。然而,人类中性粒细胞肽也能杀死正常细 胞,如成纤维细胞、上皮细胞和白细胞。此外,植物 源性防御素对肿瘤细胞也缺乏选择性[29]。因此,防御 素通常不是有效的酰基-载体蛋白质抗肿瘤药物,它们 必须进行结构优化才能实现特异性的抗肿瘤作用。
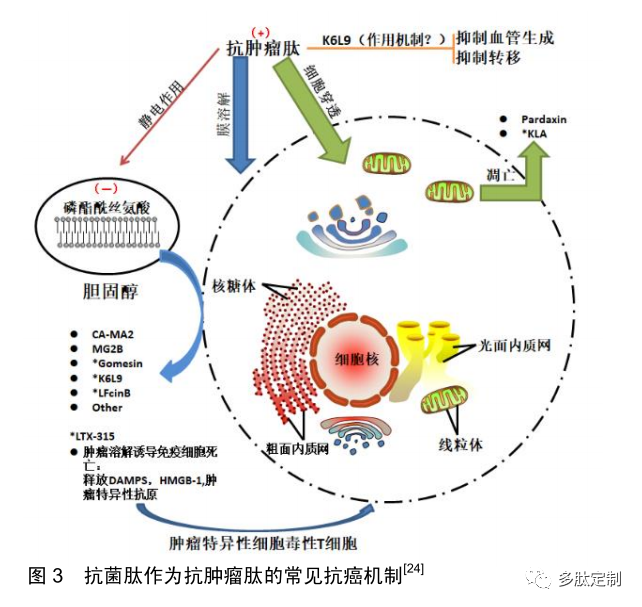
2.3.2 抗菌肽的选择性膜破坏作用 最早被发现的一 种具有抗肿瘤治疗功能的抗菌肽是Magainin,它是一类在 非洲爪蟾幼虫中发现的具有广谱抗菌活性的抗菌肽30。Magainin 2 及其类似物对血液及实体肿瘤细胞都显示出 具有选择性,12mg/L 的Magainin在 1h 内便能迅速产生肿 瘤杀伤作用;即使质量浓度升高至200 mg/L 时,其对正 常淋巴细胞仍未表现出细胞毒性。抗菌肽诱导的离子通道 作用导致钠离子、钾离子和氯离子的泄漏。如果消除细胞 膜的电离子梯度,抗菌肽的细胞毒性作用即被消除,表明膜电位对抗菌肽抗肿瘤活性至关重要。
由于抗菌肽在体内实验中没有表现出的抗肿瘤活 性,且其抗肿瘤细胞活性比抗菌活性低很多,因此还有 2个主要问题有待解决:抗肿瘤活性有限;对肿瘤细胞 的特异性低。如果抗菌肽对肿瘤细胞的特异性能够增 强,则可以克服以上2个问题。抗菌肽与肿瘤细胞的相 互作用强度可能影响其活性和特异性。因此学者们对很 多抗肿瘤肽进行结构优化来达到增强治疗效果的目的。由于天然抗菌肽在免疫系统中发挥多种作用,因此它们 在预防宿主免受疾病侵害方面比治疗方面更有效[31]。因 此,要提高治疗作用有效的做法是对抗菌肽结构进行优 化,而不能直接使用自然的抗菌肽。而合理的抗菌肽设 计需要建立一个对宿主靶细胞具有独特选择性的框架 区域,以便区分阳离子和疏水结构域,最终设计出可增 强治疗效果的合成抗菌肽。
2.3.3 抗菌肽的非膜溶解机制 研究者利用双亲性原 理设计了一个理想化的双亲螺旋结构,该抗菌肽K6L9 具 有15个残基,其对癌细胞发挥直接膜溶解作用的同时,还 显示出具有抗血管生成和抑制转移的作用,但其具体的抗 肿瘤机制还有待进一步阐明。另一种常见的抗肿瘤机制是 促进肿瘤细胞凋亡[2。尽管与正常细胞膜相比,抗肿瘤肽 对癌细胞的抑制作用具有选择性,但一些抗菌肽可能同时 扰乱细胞膜并穿透细胞,而其他非溶解性抗肿瘤肽可能仅 仅具有穿透细胞作用。当2种作用同时存在的情况下,抗 肿瘤肽能够破坏线粒体,并导致细胞程序性死亡。这种凋 亡机制可以通过线粒体肿胀、线粒体膜破裂及刺激凋亡标 记物的活化来实现。曾有学者将抗肿瘤肽注射至肿瘤内 部,使嵌合肽转移到肿瘤细胞的细胞质中,破坏软骨组织, 并通过caspase-3激活凋亡酶级联。其他抗肿瘤肽具有穿 透癌细胞的固有特性,不需要与细胞穿透肽结合。
例如pardaxin(GFFALIPKIISSPLFKTLLSAVGSALSSSGGQE)可以通过caspase-3激活诱导鳞癌细胞凋亡[3。某些抗 菌肽导致的免疫原性细胞死亡,能够对相同肿瘤产生长期 免疫保护作用。抗肿瘤肽LTX-315便具有上述特性34,其 相关应用目前已进入一期临床试验[35。
综上所述,随着抗菌肽抗癌机制的逐步阐明,通过 优化抗菌肽结构来增强抗癌效果将更容易实现,其抗肿 瘤的临床作用在未来具有广泛的应用前景。
2.4 与抗肿瘤相关的抗菌肽
2.4.1 防御素 防御素是真核生物产生的阳离子肽,包含2个超家族,在序列、结构及功能上得到了不同的水 平的改造。各种天然或合成的防御素在大量肿瘤细胞实 验或肿瘤动物模型实验中已经被证实具有良好的肿瘤 抑制作用。有研究表明,人类β-防御素3能够抑制血管 内皮生长因子诱导的癌细胞迁移能力[2。另一项研究发 现人类β-防御素3是由肿瘤浸润性单核细胞产生的,并 以剂量依赖和旁分泌的方式抑制结肠癌细胞的侵袭和 转移能力[36]。以上体外实验结果所显示出的恶性表征与 转移灶具有一致性,而防御素在抑制肿瘤细胞迁移方面 的有效性,说明其可以作为潜在的转移抑制剂。此外, 天然防御素似乎对肿瘤细胞亦可以起到抑制增殖和促 进凋亡的作用,并诱导细胞周期停滞137-38],该作用与视 网膜母细胞瘤蛋白磷酸化水平的增加,转录和细胞周期 依赖性激酶及其催化周期蛋白活性抑制, 以 及 caspase-7和caspase-9等凋亡相关性标志物表达上调 有关。除了天然衍生物,合成防御素类似物可以产生更 大的抗癌效果:有研究证明将人β-防御素2和人血清白 蛋白重组成特异性防御素后,其在抑制K-ras突变胰腺 癌MIAPaCa-2 细胞系的肿瘤增殖作用比天然β-防御素 更有效[39,见表1。
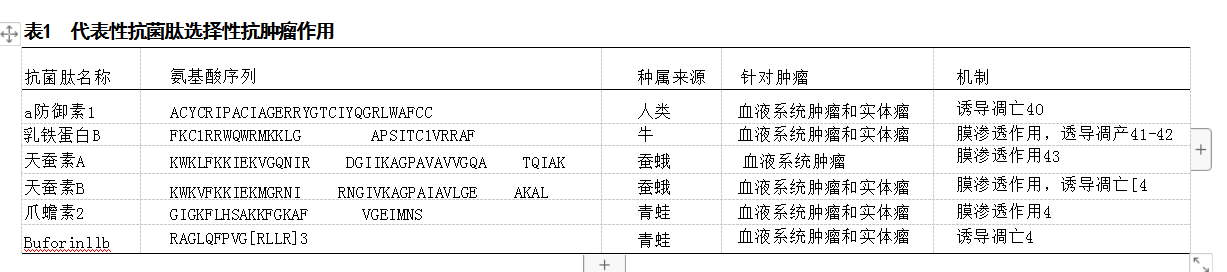
人类防御素还具有另外2个重要的特性。第一,它 们对肿瘤细胞具有显著的特异性,而对正常的细胞似乎 没有明显的细胞毒性或细胞抑制作用[37-38.47]。已有研究 表明,防御素通过内在途径诱导MCF-7 细胞凋亡,增强 MAPK p38磷酸化,增加细胞色素c、APAF-1、caspase-7 和caspase-9的表达,但不影响膜电位和钙流量[3]。这 一结果表明人类抗菌肽内源性衍生的特性,即可以避免 在炎症反应过程中对正常健康组织造成非特异性杀伤。第二,抗菌防御素可能对一些尚未解决的临床问题提供 了新的治疗机会,如化疗耐药问题。研究表明,防御素 与细胞毒性化疗药物(如阿霉素)联合应用于多重耐药肿 瘤细胞时可有效增强细胞杀伤作用3。
2.4.2 乳铁蛋白肽B 乳铁蛋白是一种分子质量80 kD 的铁隔离糖蛋白,存在于乳汁等外分泌液和多形核白细 胞颗粒中。乳铁蛋白B 具有一定的抗肿瘤作用,因为它 能够对肿瘤细胞的胞质膜和线粒体膜产生致命的、选择 性的失稳作用[41,并且已被证明对结直肠癌、神经母细 胞瘤和黑色素瘤癌细胞有效42.48-49。抗菌肽多具有多种 构象,最近一项研究表明,牛乳铁蛋白(bLf)、环LfcinB 和线性LfcinB均能激活多种抗肿瘤信号级联,通过转录组学分析其能够促进p53激活、凋亡发生以及抗血管生 成信号通路的活化。此外, bLf和LfcinB均诱导了 caspase-8、p53和p21介导的大肠癌细胞凋亡前通路42。与防御素一样,乳铁蛋白似乎具有免疫调节作用,并已 被证明能使淋巴细胞定向根除癌细胞501。
2.4.3 天蚕素A和B 天蚕素是一类典型的短碱性肽,长 度在31-39个氨基酸残基之间,具有一个强碱性的N 末端 和疏水性的C 末端,其最初从天蚕中分离出来的,其抗癌 机制和其他抗菌肽相类似,天蚕素A和B被证明对膀胱肿 瘤有选择性的细胞毒性和细胞抑制作用,但对人类成纤 维细胞株没有作用43],而ABP-dHC- 天蚕素A 及其类似物 ABP-dHC- 天蚕素A-K对白血病细胞株有细胞毒性,但对 正常体细胞株没有毒性作用[44。天蚕素A 通过ROS 信号 传导通路在早幼粒细胞系HL-60 中诱导凋亡144,并通过 Fas、Fas-I、caspase-3和caspase-8的表达在人肝癌细 胞中诱导凋亡51。天蚕素XJ通过caspase-3和多聚(ADP- 核糖)多聚酶的表达及对Bcl-2的下调诱导肝细胞癌细胞 的生长抑制、 S 期阻滞和凋亡145。在体外实验中,天蚕素 A增强了常用化疗药物对鳞状皮肤癌细胞的作用,这也为 该肿瘤提供了新的治疗策略。
2.4.4 爪蟾素Ⅱ 爪蟾素Ⅱ是一种从非洲爪蛙的皮肤 中分离出来的抗菌肽,在严重的造血和实体恶性肿瘤中 显示出有效的抗癌作用。作为一种具有螺旋结构的离子 载体肽,它已被证明能够穿透癌细胞膜,起到离子通道 的作用,导致癌细胞的细胞溶解[521,从而显著增强了细 胞毒性化疗药物的抗肿瘤作用。例如,当与治疗非小细 胞肺癌细胞的化疗药物一起使用时,爪蟾素A和G 具有协 同作用。爪蟾素Ⅱ也表现出抗肿瘤的选择性,例如,它 们已被证明能溶解各种造血肿瘤和实体肿瘤细胞,但对 正常人成纤维细胞和外周血淋巴细胞几乎没有影响[15]。同时,爪蟾素Ⅱ的衍生物可以增强对肿瘤细胞的特异性细胞毒性521。
2.4.5 其他具有抗肿瘤特性的代表性抗菌肽 在结构 上,LTX-302 是一种源于其亲本肽LfcinB的9-聚体肽, 具有优化的α-螺旋二级结构,从而发挥巨大的抗肿瘤活 性。LTX-302在BALB/c小鼠体内建立的A2O-B 细胞淋巴 瘤模型中已被证实具有明显的抗肿瘤作用,其抗肿瘤活 性依赖于CD4 和CD8 淋巴T 细胞的活化,且最终LTX-302 给药诱导了针对淋巴瘤细胞的长效细胞免疫,最终导致 肿瘤消退[14。Buforin II是一种具有21残基的α-螺旋抗菌 肽,其序列与组蛋白H2A 的N 末端区域相似。Buforin IIB 是一种合成衍生的类似于Buforin II的化合物,经修饰后 在2个a-螺旋之间含有脯氨酸残基153]。在一项研究中发 现Buforin IIB对乳腺癌细胞MX-1 和MCF-7 具有抗癌活 性,并通过抗血管生成机制抑制小鼠动物模型中种植瘤 的生长15。由于此次综述篇幅有限,在该部分仅涉及到 以上具有代表性的抗菌肽。
2.5 抗菌肽的临床试验 一些阳离子抗菌肽可能主要通 过诱导宿主免疫反应而不是直接作用方式发挥其杀菌 作用。因此在过去几年中,它们作为抗癌药物的辅助药 物而被广泛研究,同时近年来肿瘤免疫治疗相关临床试 验也证实了这一作用机制。
商业上开发的第一个抗菌肽是乙酸培昔甘南 (MS1-78)⁴6。此外,已经有很多抗菌肽正在系统研发中。例如, 一种正在处于临床试验阶段的抗菌肽是omiganan, 它是从牛中性粒细胞中纯化的吲哚菌素衍生物,它以凝胶 的方式被用作预防中心静脉导管污染[54。此外,与氧氟沙 星相比,吡昔甘南可以作为治疗糖尿病患者轻度感染足部 溃疡的有效替代品155。
目前抗菌肽刚开始应用于肿瘤领域,其疗效数据相对有限。然而,从既往感染相关性临床试验中所得到的安全性数据,间接支持抗菌肽将在癌症患者中具有较好 的耐受性。正在进行的肿瘤临床试验是NCTO225366,其中LL37 对转移性黑色素瘤的最佳生物剂量和治疗有 效性正在临床一期评估中。LL37通过瘤内注射的方式来 治疗转移性黑色素瘤患者。
3 展望 Prospects
抗菌肽作为抗肿瘤肽的临床发展与抗菌药物的临 床发展面临着同样的挑战。抗菌肽不是传统的抗菌药, 因其具有多功能特性,因此在不进行结构优化的情况 下将其直接应用于临床是相当具有挑战性的。虽然抗菌肽在免疫系统中工作良好,但在本质上, 一个特定 的抗菌肽结构可能没有针对单个功能(如抗菌、抗病毒 或抗癌)进行完全优化。除了抗菌肽的稳定性之外,另 一个重要的关注点在于体外敏感性试验与动物模型之间缺乏有效的相关性,在动物模型中建立体外稳定性和生物利用度之间相关性的分析可能解决这一问题。 目前,结构优化后的抗菌肽可以实现非肠道途径给药, 其中包括经静脉途径、呼吸道途径、肌肉注射、腹膜内及皮下给药[56-58]。尽管抗菌肽的理化性质阻止其经 口服用,但开发特定的输送系统保护抗菌肽不被消化 酶降解,可能会增加肠道吸收和口服的可行性。此外, 应用非人类分泌的合成抗菌肽,其毒性仍然是一个突出的问题。然而可以从传染病领域汲取经验,在该领域数个抗菌肽已过渡到临床试验,甚至在临床应用中 获得了有效证据。例如可以通过开发创新配方和药物 输送系统来提高这些肽治疗指数。结构-活性关系也对 提高抗菌肽的抗肿瘤效果至关重要。当然, 目前这些 抗肿瘤机制还仍是推测。此外, 自然界中存在的绝大 多数抗菌肽尚未被特异分离出来,考虑到它们在治疗 应用方面的潜力,更全面的研究仍很有必要。未来以 合理的成本将这些抗菌肽应用于抗肿瘤治疗中,将使这一新兴治疗方法具有光明的应用前景。
免责声明:本文为行业交流学习,版权归原作者所有,如有侵权,可联系删除。